Сравнение эффективности компактных УФ-ламп для синтеза витамина D в коже растущих бородатых агам
J. J. E. Diehl, F. M. Baines, A. C. Heijboer, J. P. van Leeuwen, M. Kik, W. H. Hendriks, D. G. A. B. Oonincx
Было изучено влияние излучения различных компактных ультрафиолетовых ламп на уровень витамина D у растущих бородатых агам (Pogona vitticeps). Сорок две недавно вылупившиеся бородатые агамы (в возрасте не старше 24 часов) были разделены на шесть групп (n=7 в каждой группе). Пять групп подвергались облучению различными компактными УФ-ламп в течение двух часов в день, а контрольная группа не подвергалась воздействию УФ-излучения. В возрасте 120 дней у них взяли образцы крови и определили концентрации 25-(OH)D3 (25-гидроксихолекальциферол, метаболит витамина D3), Ca, P и мочевой кислоты. Кроме того, определили концентрацию 25-(OH)D3 в плазме у живущих в природе взрослых бородатых агам для установления контрольного уровня. Только в одной экспериментальной группе уровень 25-(ОН)D3 был повышен по сравнению с контрольной группой (41,0 ± 12,85 против 2,0 ± 0,0 нмоль/л). У всех групп, облучаемых ультрафиолетом, были низкие уровни 25-(OH)D3 в плазме по сравнению с более ранними исследованиями бородатых агам в неволе, а также по сравнению со взрослыми бородатыми агамами, живущими в природе (409 ± 56 нмоль/л). Спектральный анализ показал, что все использованные в исследовании лампы излучали UVB, эффективный для синтеза витамина D в коже. Ни одна из этих ламп в таком режиме, по-видимому, не обеспечивала дозу UVB-излучения, достаточную для синтеза такого же уровня 25-(OH)D3 в плазме, как у диких бородатых агам в их естественной среде обитания.
1. Введение
Бородатая агама (Pogona vitticeps) – один из наиболее популярных видов среди владельцев рептилий в Северной Америке (Wright, 2008) и, возможно, в Европе. Наиболее частые патологии, которые возникают при содержании P. vitticeps, относятся к группе метаболических заболеваний костей (MBD) (Wright, 2008). Этот комплекс заболеваний включает рахит, остеопороз и вторичный гиперпаратиреоз и вызван дисбалансом кальция и/или фосфора в организме (Divers & Mader, 2005). Витамин D3 регулирует метаболизм кальция, способствуя кишечной абсорбции (всасывание) и почечной реабсорбции (обратное всасывание) Ca и P (Ajibade, Benn & Christakos, 2010; Haxhiu et al., 2014). Кроме того, дисбаланс в рационе витамина D, Ca или P может вызвать заболевание почек у ящериц (Miller, 1998), что приводит к повышению уровня фосфора и мочевой кислоты в плазме (Knotek, Hauptman, Knotkova, Hajkova, & Tichý, 2002).
Дефицит витамина D3 может привести к гипокальциемии, которая, в свою очередь, может привести к развитию MBD (Holick, Tian, & Allen, 1995). Витамин D3 может быть либо получен экзогенно, либо эндогенно синтезирован в коже. Эпидермальные клетки кожи содержат 7-дегидрохолестерин (7-DHC), который превращается в пре-витамин D3 при облучении ультрафиолетом B в диапазоне длин волн 290-320нм (Fraser, 1995). Пре-витамин D3 изомеризуется в витамин D3 на температура-зависимой стадии (Holick et al., 1980). Витамин D3 в коже связывается с витамин D-связывающим белком (VDBP), который транспортирует его в кровоток. Витамин D3 гидроксилируется в печени до 25-(OH)D3, который снова связывается с VDBP и возвращается в кровь. Циркулирующий 25-(OH)D3 считается основной формой хранения витамина D3 и используется для оценки уровня витамина D3 (Gillespie, Frye, Stockham, Fredeking, 2000 ; Holick, 1990). В клетках почек 25-(OH)D3 переходит в активный эндокринный гормон 1,25-(OH)2D3 (1,25-дигидроксихолекальциферол), который является жизненно важным регулятором гомеостаза кальция и фосфора. У людей и, возможно, у других позвоночных, 25-(OH)D3 также попадает в клетки многих других органов, в которых есть рецепторы витамина D3, включая иммунную систему, где происходит внутриклеточное превращение в 1,25-(OH)2D3 (Hossein-nezhad & Holick, 2013). В клетках 1,25-(OH)2D3 выполняет паракринную и аутокринную функции (Björn, 2008).
Витамин D3 используют почти все изученные виды позвоночных (Björn, 2008), и синтез витамина D3 из 7-DHC под действием ультрафиолета – древний процесс, который сохранился в ходе эволюции. Большинство позвоночных животных могут использовать уже готовый витамин D3 из пищи, что позволило содержать в неволе многие виды рептилий без облучения UVB, но с добавлением витамина D3 в рацион. Первым успешное использование флуоресцентной трубки, излучающей ультрафиолет B, для синтеза витамина D3 у рептилий описал Ласло в 1969 году.
С тех пор синтез витамина D3 в коже под действием искусственного UVB-излучения был продемонстрирован у нескольких дневных рептилий (Acierno, Mitchell, Roundtree, & Zachariah, 2006; Allen, Oftedal, & Horst, 1995; Bernard, Allen, & Ullrey, 1997; Ferguson et al., 2003 , 2009; Gillespie et al., 2000; Hibma, 2004; Hoby et al., 2010; Kroenlein, Zimmerman, Saunders & Holladay, 2011; Oonincx, Stevens, van den Borne, van Leeuwen, & Hendriks, 2010; Selleri & Di Girolamo, 2012), а также у сумеречных и ночных видов (Acierno et al., 2008; Carman, Ferguson, Gehrmann, Chen, & Holick, 2000 ; Wangen, Kirchenbaum, & Mitchell, 2013). Большинство из этих исследований не описывают интенсивность UVB и спектральное распределение, полученные испытуемыми. Спектральное распределение мощности источника света определяет полученные фото продукты (MacLaughlin, Anderson, & Holick, 1982). Состав люминофора и пропускная способность внешней стеклянной трубки люминесцентной лампы, излучающей UVB, определяют спектральное распределение мощности и, следовательно, ее потенциал относительно синтеза витамина D3. Кроме того, виды животных, вероятно, будут иметь разные оптимальные уровни воздействия ультрафиолета B из-за различий в среде обитания и в характере баскинга (прогрев под солнцем и получение УФ) (Ferguson et al., 2005) и могут также различаться в способности усваивать витамин D3, поступающий с пищей (Allen et al., 1995). Эти различия и отсутствие эталонных значений для концентрации 25-(OH)D3 в плазме у живущих в природе особей большинства видов затрудняют определение «адекватных» уровней UVB-излучения для рептилий в неволе. Контрольные значения необходимы для определения эффективности любого режима приема добавок, будь то алиментарный путь (поступает с пищей) или облучение UVB. Концентрации метаболитов витамина D у живущих в природе рептилий были зарегистрированы только для комодского варана (Gillespie et al., 2000), чаквеллы Sauromalus obesus (Aucone, Gehrmann, Ferguson, Chen & Holick, 2003), циклуры Рикорда Cyclura ricordii и игуаны-носорога Cyclura cornuta cornuta (Ramer et al., 2005).
В этом исследовании оценивается влияние длительного воздействия пяти компактных УФ-ламп, продаваемых для содержания рептилий, на концентрацию 25-(OH)D3 в плазме бородатых агам. Мы предполагаем, что спектральное распределение мощности этих ламп определяет уровень витамина D у облучаемых бородатых агам. Кроме того, для оценки этих уровней 25-(OH)D3 в плазме были взяты образцы крови диких живущих в природе бородатых агам в Австралии и проанализированы на содержание 25-(OH)D3.
2. Материалы и методы
2.1 Испытание и животные
Это исследование было одобрено Комитетом по уходу и использованию животных Университета Вагенингена, Нидерланды. Исследование состояло из шести параллельных испытаний: пять типов коммерческих компактных люминесцентных УФ-ламп и одна контрольная лампа, которая не излучала UVB. Всего 42 особи (19 самцов и 23 самки) из двух кладок недавно вылупившихся бородатых агам (P. vitticeps), отложенных одной парой, были получены от частного заводчика. Начиная со дня вылупления, еженедельно измеряли длину от конца морды до клоакальной щели (SVL) и общую длину (TL) и определяли массу тела (BM), используя точные весы (тип HF 2000G, A&D Company Ltd, Токио, Япония). Бородатых агам распределили в шесть террариумов в течение 24 часов после вылупления. Чтобы уменьшить разницу в возрасте, с 1 по 7 вылупившихся животных распределили в первый террариум, с 8 по 14 – во второй террариум и так далее, пока все террариумы не содержали по семь животных.
2.2 Конструкция и размеры
В каждый террариум с размерами 124×50×40см (Д×Ш×В) установили 60-ваттную зеркальную лампу (лампа Philips NR80; Philips Eindhoven, Нидерланды), которая ежедневно обеспечивала тепло с 08:00 до 22:00, и компактную люминесцентную лампу. В этом исследовании использовали одну контрольную компактную люминесцентную лампу без UVB-излучения и пять компактных люминесцентных ламп, продаваемых для содержания «пустынных» рептилий (таблица 1). Перед началом исследования лампы использовались в течение 100 часов для стабилизации выхода UVB. Когда первое животное было помещено в террариум, компактные люминесцентные лампы включались с 12:00 до 14:00 ежедневно с использованием таймера.
| БРЕНД | ТИП ЛАМПЫ | МОЩНОСТЬ (Вт) |
|---|---|---|
| Control | Philips SoftoneT65 WW827 | 20 |
| Trixie | Trixie Desert Pro Compact 10.0 | 23 |
| JBL | JBL Reptile Desert UV 480 Desert Terrarium Lamp | 23 |
| Arcadia | Arcadia D3 + 10%UVB Compact Reptile Lamp | 23 |
| ZooMed | ZooMed ReptiSun 10.0 UVB Desert Compact Fluorescent Lamp | 26 |
| ExoTerra | ExoTerra Repti Glo 10.0 UVB Desert Terrarium Lamp | 26 |
Один кирпич (21×10×5см) поместили под зеркальную лампу, а другой – непосредственно под компактную люминесцентную лампу, чтобы организовать место для баскинга высотой 5см (рис. 1). Зеркальная лампа располагалась на 15см выше места для баскинга, а компактная люминесцентная лампа была установлена горизонтально на высоте 25см над вторым местом для баскинга. Уровень UVB измеряли два раза в неделю в двух точках террариума: А) непосредственно под центром компактной УФ-лампы на расстоянии 21,5см и В) в передней части террариума возле стекла, на расстоянии 31см от центра лампы (рис. 1). Общий выход UVB (мкВт/см²) и индекс УФ UVI определяли с помощью радиометров (Solartech Inc., Harrison Township, MI, USA; модели 6.2 и 6.5).
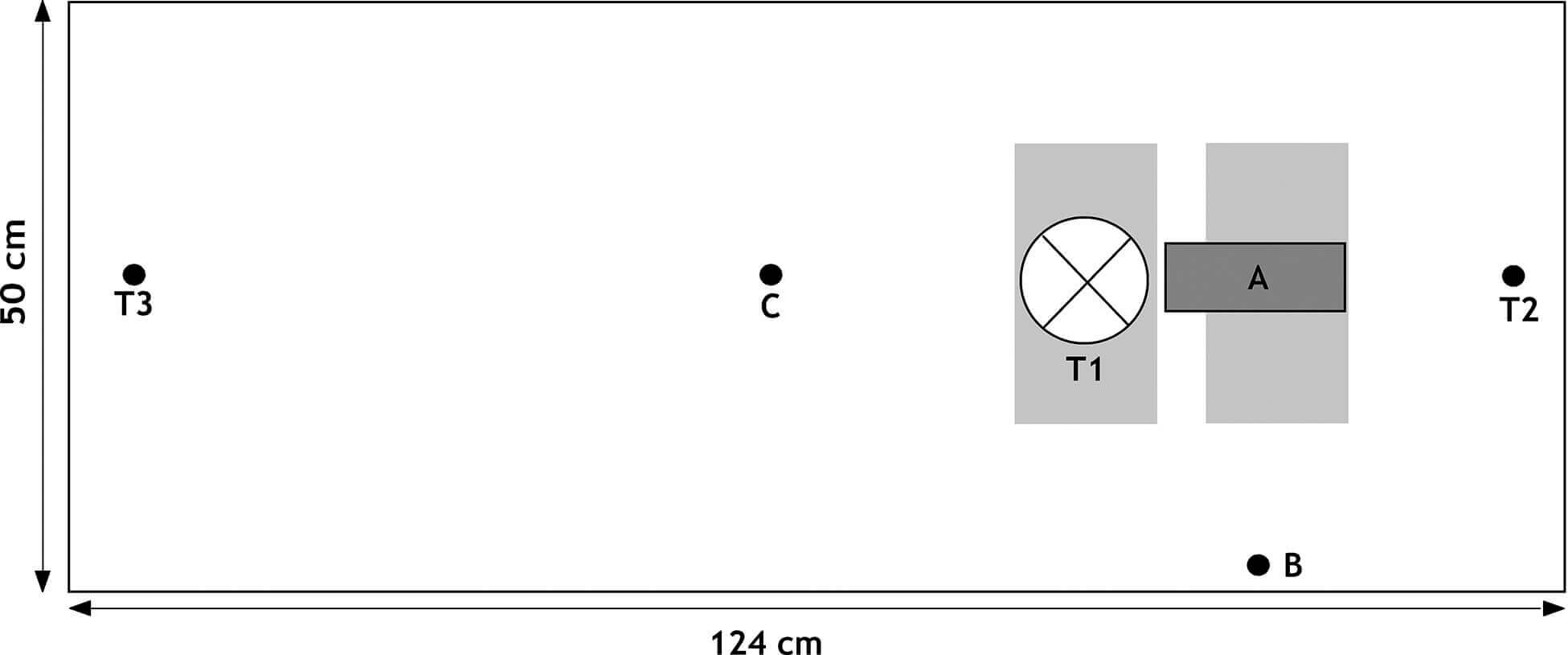
Расположение террариума: A и B указывают, где измеряли интенсивность UVB и индекс УФ UVI; T1, T2, T3 – точки измерения температуры; зеркальная лампа; УФ-лампа; кирпич.
Каждую неделю перед включением компактных ламп температуру поверхности измеряли непосредственно под зеркальной лампой (T1, рисунок 1) и в 3см с каждой стороны террариума (T2 и T3, рисунок 1) с использованием инфракрасного термометра RayTemp 3 (Electronic Temperature Instruments Ltd, West Sussex, UK).
В возрасте 120 дней из вентральной хвостовой вены каждого животного взяли образец крови (1,5 мл) с использованием гепаринизированного шприца объемом 2мл, содержащего 80МЕ электролитно-сбалансированного гепарина (шприц PICO50, Radiometer Medical ApS, Brønshøj, Дания) и иглы 25-го калибра (BD Microlance 3 25G 0,5 × 16 мм ref 300600). Сразу после сбора образец разделили на две равные части, одну часть центрифугировали (750×g в течение 10 минут при 10°C) и хранили при -80°C в течение 1 часа после экстракции до анализа на 25-(OH)D3. Вторую половину хранили в закрытом шприце и отправили на анализ для определения концентрации кальция, фосфора и мочевой кислоты.
2.3 Рацион и кормление
На протяжении всего исследования бородатые агамы получали ad libitum (по желанию) домашних сверчков (Acheta domesticus), пустынную саранчу (Schistocerca gregaria), азиатскую саранчу (Locusta migratoria) и мучного хрущака (Tenebrio molitor) подходящего размера в соответствии с их массой тела. Все насекомые были приобретены у коммерческого поставщика (Star Food BV, Barneveld, Нидерланды) и перед подачей посыпались порошком карбоната кальция. Потребление корма по пищевому составу определяли ежедневно на террариум и рассчитывали путем вычитания веса отказов от веса, предложенного накануне. В конце исследования среднее общее потребление корма рассчитывали по пищевому составу.
2.4 Образцы крови живущих в природе бородатых агам
Разрешение на научные исследования было получено в Службе национальных парков и диких животных Департамента охраны окружающей среды и охраны природы штата Нью-Йорк в соответствии с Законом о национальных парках и дикой природе 1974 года, раздел 132C. Кроме того, разрешение в соответствии с Законом об исследованиях на животных было выдано Отделом защиты животных Министерства сельского хозяйства и инвестиций штата Новый Южный Уэльс (номер лицензии S13043). Бородатые агамы были пойманы вручную в Новом Южном Уэльсе, между городами Брокен-Хилл (S 31.91021°, E 141.48497°) и Кобар (S 31.71187°, E 144.15192°) летом 2010 года (28 января-8 февраля) (Oonincx, van Leeuwen, Hendriks, van der Poel, 2015). Пол определяли с помощью визуального осмотра. Образцы крови у тринадцати взрослых (четыре самки и девять самцов) и у одного подростка мужского пола были взяты, как описано выше. Эти образцы крови центрифугировали в местных больницах в течение 12 часов и хранили в мобильном морозильнике (Engel Fridge, Engel, Australia), а затем отправили на сухом льду в Университет Вагенингена, Вагенинген, Нидерланды. Во время каждой поимки на данном месте фиксировали показания солнечного UVB-излучения с использованием UVB-метра Solarmeter модели 6.2.
2.5 Лабораторные анализы
Образцы крови бородатых агам в эксперименте с УФ-лампой были проанализированы на содержание 25-(OH)D3 в плазме крови в эндокринной лаборатории Медицинского центра университета VU (Амстердам, Нидерланды) с помощью ID-XLC-MS/MS, как описали Heijboer, Blankenstein, Kema, Buijs (2012) с модификациями для оптимизации метода для бородатых агам, как описал Oonincx et al. (2013). Для проведения статистического анализа пробам ниже предела обнаружения (4нмоль/л) присваивали произвольное значение 2нмоль/л. Расчеты с 0, 2 или 4нмоль/л не влияли на результаты статистического анализа. Плазменные концентрации метаболитов витамина D (25-(ОН)D3 и 1,25-(ОН)2D3) у живущих в природе бородатых агам определяли индивидуально радиоиммуноанализом, как описал Oonincx et al. (2010).
Общие концентрации Ca, P и мочевой кислоты (UA) определяли с помощью прибора Unicel DXC-600 (Beckman Coulter, Woerden, Нидерланды). Концентрацию ионизированного Са определяли газоанализатором крови (Rapidlab 1265, Siemens Nederland BV, Гаага, Нидерланды) в Департаменте клинических наук животных-компаньонов Утрехтского университета (Утрехт, Нидерланды).
2.6 Оценка ламп
В конце испытания люминесцентные лампы, использованные в исследовании, были установлены на испытательном стенде, и после стандартного 30-минутного разогрева на сетевом напряжении, регулируемом с помощью трансформатора Variac (Carroll and Meynell CMV2E-1) до 230В, показания счетчиков снимались с каждой лампы с помощью прибора для измерения UVI Solarmeter 6.5 и широкополосного измерителя UVB Solarmeter 6.2. Записи были сделаны со стороны каждой лампы, при этом датчик располагался перпендикулярно оси лампы, на полпути вниз спиральной части колбы. Показания счетчика снимались с шагом 5см от поверхности лампы.
Спектрограммы были получены с использованием спектрального радиометра Ocean Optics USB2000+ с UVB-совместимым оптоволоконным зондом с косинусным адаптером (Ocean Optics Inc., Данидин, Флорида, США). Спектрометрические записи производились на стандартном расстоянии 10см от поверхности лампы. Измеренное спектральное излучение (мкВт/см2/нм) от каждой лампы сравнили с эталонным CIE спектром, активным для превитамина D3 (Bouillon et al., 2006), чтобы получить сравнение их витамин D-синтезирующего излучения (D-Eff). В предыдущем эксперименте (Oonincx et al., 2010) использовали аналогичные экспериментальные параметры другой компактной лампы. Было проведено сравнение со спектром и D-Eff этой лампы (ZooMed Reptisun 5.0 UVb Tropical 26-ватт, ZooMed Laboratories Inc., Сан-Луис-Обиспо, США) и с эталонными солнечными спектрами из Австралии (Bernhard, Mayer, Seckmeyer, Moise, 1997).
2.7 Статистический анализ
Отдельные ящерицы считались экспериментальными единицами; взаимовлияние отдельных ящериц в террариуме, как предполагалось, было незначительным, но не могло быть оценено из-за плана эксперимента. Чтобы проверить, происходили ли животные из однородной популяции, инкубационные BM, SVL и TL были подвергнуты дисперсионному анализу облучения, пола и взаимодействия между облучением и полом как факторами. Дисперсионный анализ облучения, пола и их взаимодействия как факторов также проводился на увеличение BM, SVL и TL приблизительно через 4 месяца. Достоверность различий (p <0,05) в среднем суточном потреблении корма, УФ-индексе (UVI), UVB и температуре между обработками проверялась однофакторным дисперсионным анализом с последующим анализом LSD post hoc. Концентрации Ca, P, UA и 25-(OH)D3 анализировали с помощью непараметрического дисперсионного анализа (критерий Краскела-Уоллиса), за которым следовал U-критерий Манна-Уитни, скорректированный для множественных сравнений. Статистический анализ проводился с использованием IBM SPSS Statistics 20 (IBM Corporation, Армонк, Нью-Йорк, США).
3. Результаты
3.1 Животные и жилищные условия
Девять животных не завершили исследование; в каждой из групп, получающих UVB-излучение, одно животное удалили из-за различий в размерах с его террариумными партнерами. Пять из них изъяли между двенадцатью и пятнадцатью неделями после начала эксперимента. Одно животное из группы ZooMed удалили в возрасте 4 недель и подвергли эвтаназии из-за серьезных травм, нанесенных террариумными партнерами. В контрольной группе трое животных (одно в возрасте 2 недель, два других в возрасте 21 недели) были удалены из-за клинических признаков дефицита кальция (тетания). Оставшиеся 33 бородатых агамы были визуально здоровыми. Данные удаленных животных были исключены из статистического анализа, за исключением потребления корма, так как это было определено для экспериментальной группы. Кровь была получена от всех животных, кроме одного из группы ZooMed. Начальные значения BM, SVL и TL бородатых агам были одинаковыми для экспериментальных групп. В конце исследования BM (p <.01), SVL (p <.02) и TL (p <.02) в контрольной группе были ниже, чем в других группах (рисунок 2). Пол не повлиял на эти параметры. Средние температуры были одинаковыми в контрольной и в экспериментальных группах в течение периода исследования, однако они различались в разных местах: непосредственно под лампой обогрева (T1) 52,2±1,7°C, с правой стороны (T2) 33,5±1,4°C и с левой стороны (T3) 29,8±1,4°C.
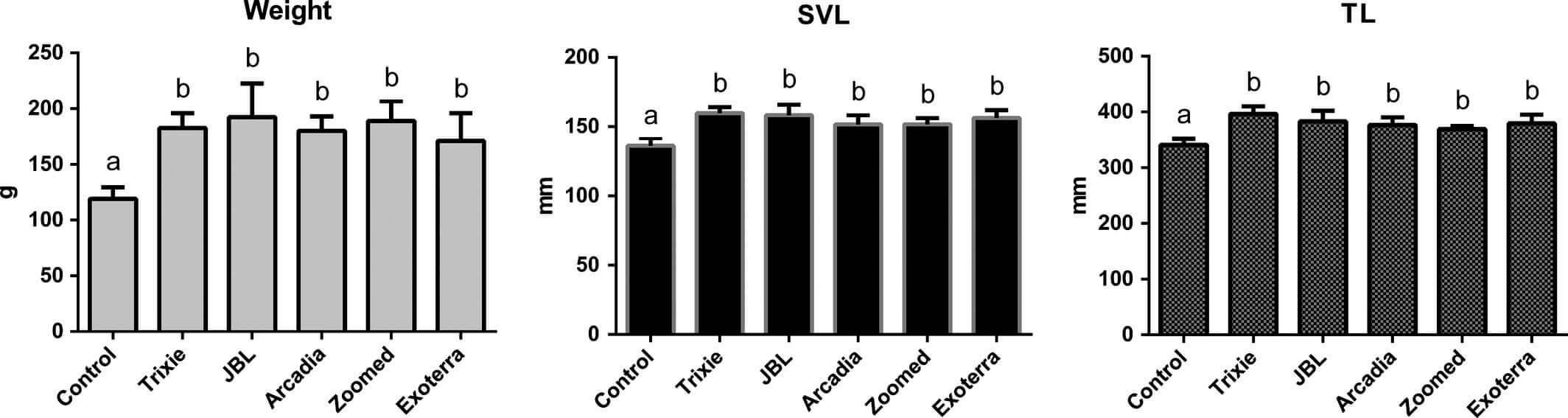
Масса тела (BM), длина от кончика морды до клоакального отверстия (SVL) и общая длина (TL) бородатых агам (Pogona vitticeps) в возрасте примерно 4 месяцев, облучаемых одной контрольной лампой и пятью компактными УФ-лампами. Различные буквы над столбцами указывают на существенные различия (p <.05)
3.2 UVB-излучение
Интенсивность UVB-излучения для пяти тестируемых ламп измеряли на разных расстояниях (таблица 2). Лампа Arcadia выдавала самое высокое UVB-излучение и UVI во всех местах; у лампы ExoTerra отмечено самое низкое UVB-излучение. UVB-излучение всех тестируемых ламп снизилось во время эксперимента (рисунок 3). Максимальное снижение было отмечено у лампы Trixie (27%), тогда как у лампы Arcadia было минимальное снижение (10%).
Таблица 2. Интенсивность UVB и УФ-индекc UVI компактных ламп в конце эксперимента, измеренные перпендикулярно лампе на разных расстояниях
| ЗНАЧЕНИЯ UVB (μW/cm2) | UV Index | |||||||
|---|---|---|---|---|---|---|---|---|
| Дистанция (см) | 20 | 25 | 30 | 35 | 20 | 25 | 30 | 35 |
| JBL | 31 | 21 | 15 | 12 | 0.9 | 0.6 | 0.4 | 0.3 |
| ZooMed | 42 | 27 | 19 | 15 | 1.4 | 0.9 | 0.7 | 0.5 |
| Arcadia | 86 | 57 | 42 | 31 | 2.9 | 1.9 | 1.4 | 1.0 |
| ExoTerra | 15 | 10 | 7 | 5 | 0.8 | 0.5 | 0.4 | 0.3 |
| Trixie | 38 | 26 | 19 | 14 | 1.1 | 0.7 | 0.5 | 0.4 |
| Control | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
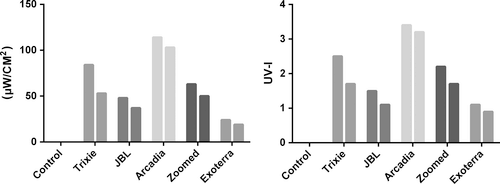
Интенсивность UVB-излучения (слева) и UVI (справа) тестируемых ламп в начале (100 часов) и в конце эксперимента (340 часов) на расстоянии 25см.
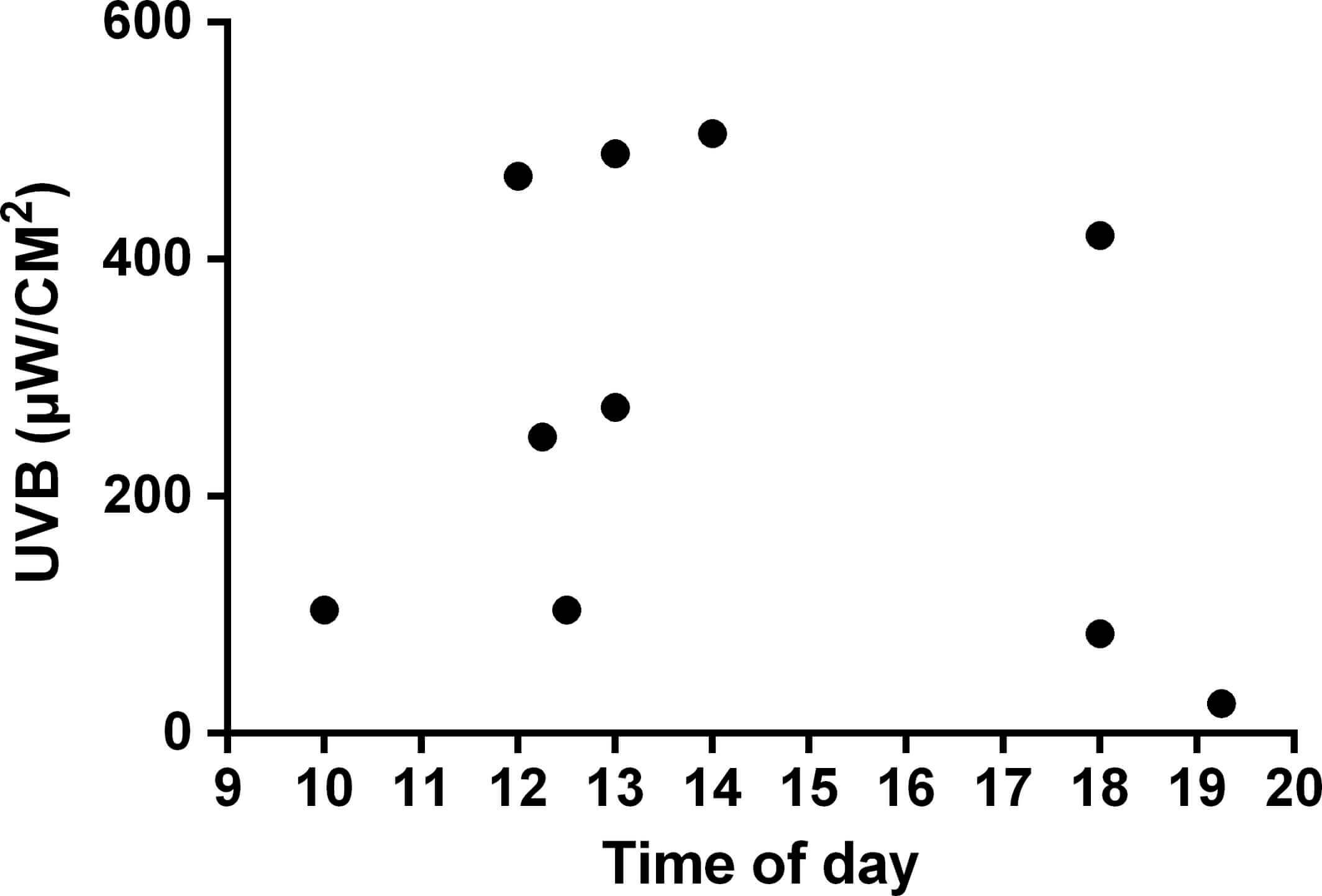
Интенсивность UVB-излучения также была определена для живущих в природе бородатых агам. Показания были взяты сразу после поимки животных в тех местах, где бородатые агамы были найдены в поле (Рисунок 4). Представленные данные включают измерения как на солнце, так и в тени. Животные часто грелись в солнечные дни, в том числе во время дневной жары. Максимальное солнечное UVB-излучение, зарегистрированное в месте расположения одной из греющихся агам, составило 506 мкВт/см2 в 14.00.
Спектры UVB-излучения каждой из пяти тестируемых ламп и лампы ZooMed из исследования Oonincx et al. (2010), далее называемый компактной лампой ZooMed 2010, показаны на рисунке 5. Спектральное распределение мощности (SPD) всех пяти тестируемых ламп было одинаковым, за исключением небольшого пика около 297 нм у лампы Arcadia. У этой лампы UVB-излучение было выше, чем у других тестируемых ламп на всех длинах волн. Компактная лампа ZooMed 2010 имела другой SPD, излучая уже на коротких длинах волн от 285 нм.
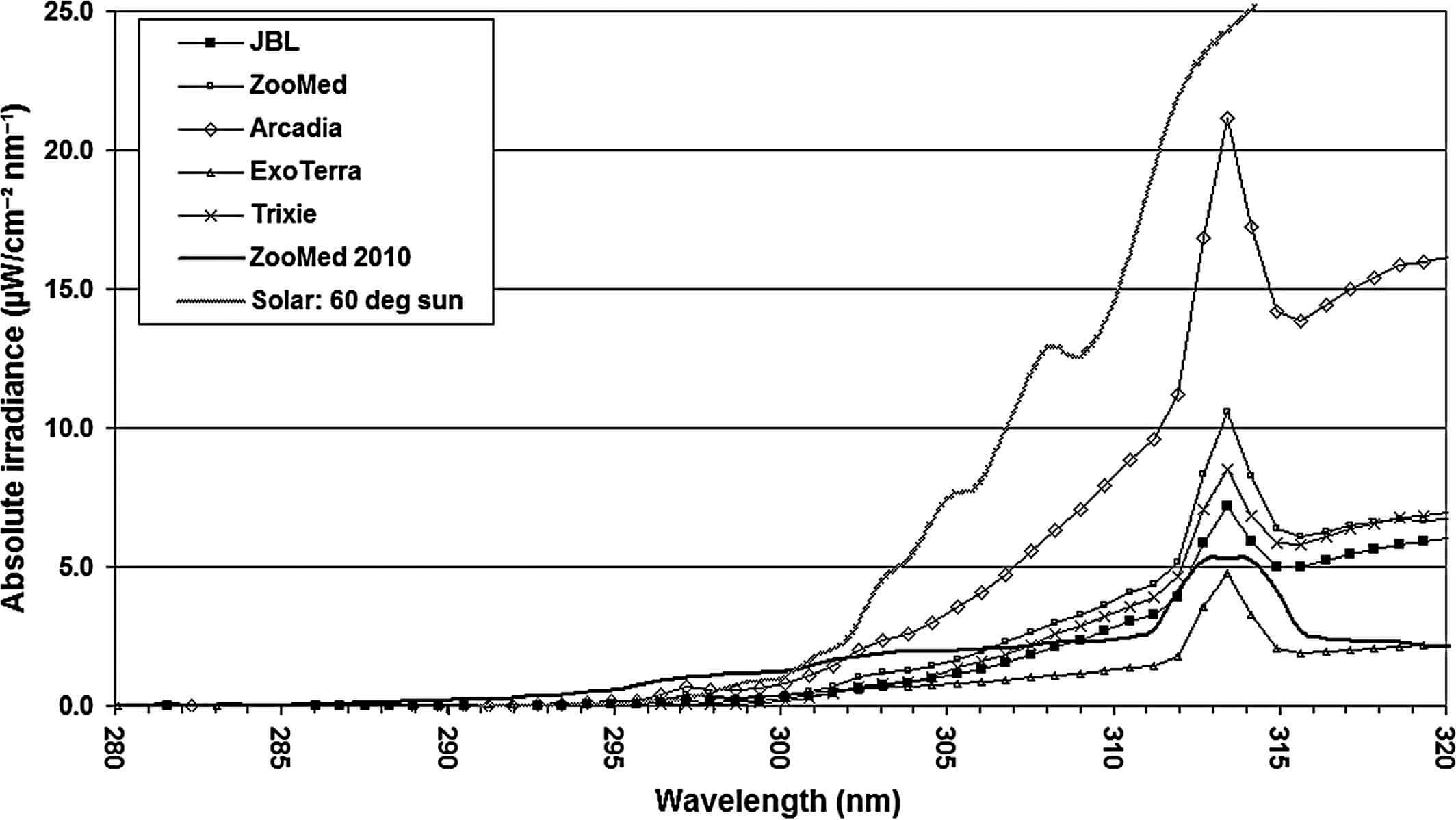
Интенсивность UVB-излучения шести компактных УФ-ламп и эталонного солнечного спектра с высотой солнца 60° (в середине утра и в середине дня летом) в Северном Квинсленде, Австралия (Bernhard et al., 1997)
Спектры D-Eff-излучений для всех ламп и солнечного света на высоте солнца 60° (Bernhard et al., 1997) показаны на рисунке 6. Общие D-Eff-излучения на расстоянии 10см, рассчитанные по спектральным данным, имеют следующий порядок исходя из их величин (мкВт/см2 D-Eff): Arcadia 28,3; ZooMed 2010 20,7; ZooMed 13,49; Trixie 10,05; JBL 9,35; ExoTerra 7.63. Солнечный свет имел намного более высокое общее D-Eff (49,5 мкВт/см2 D-Eff), чем у всех ламп на расстоянии 10см.
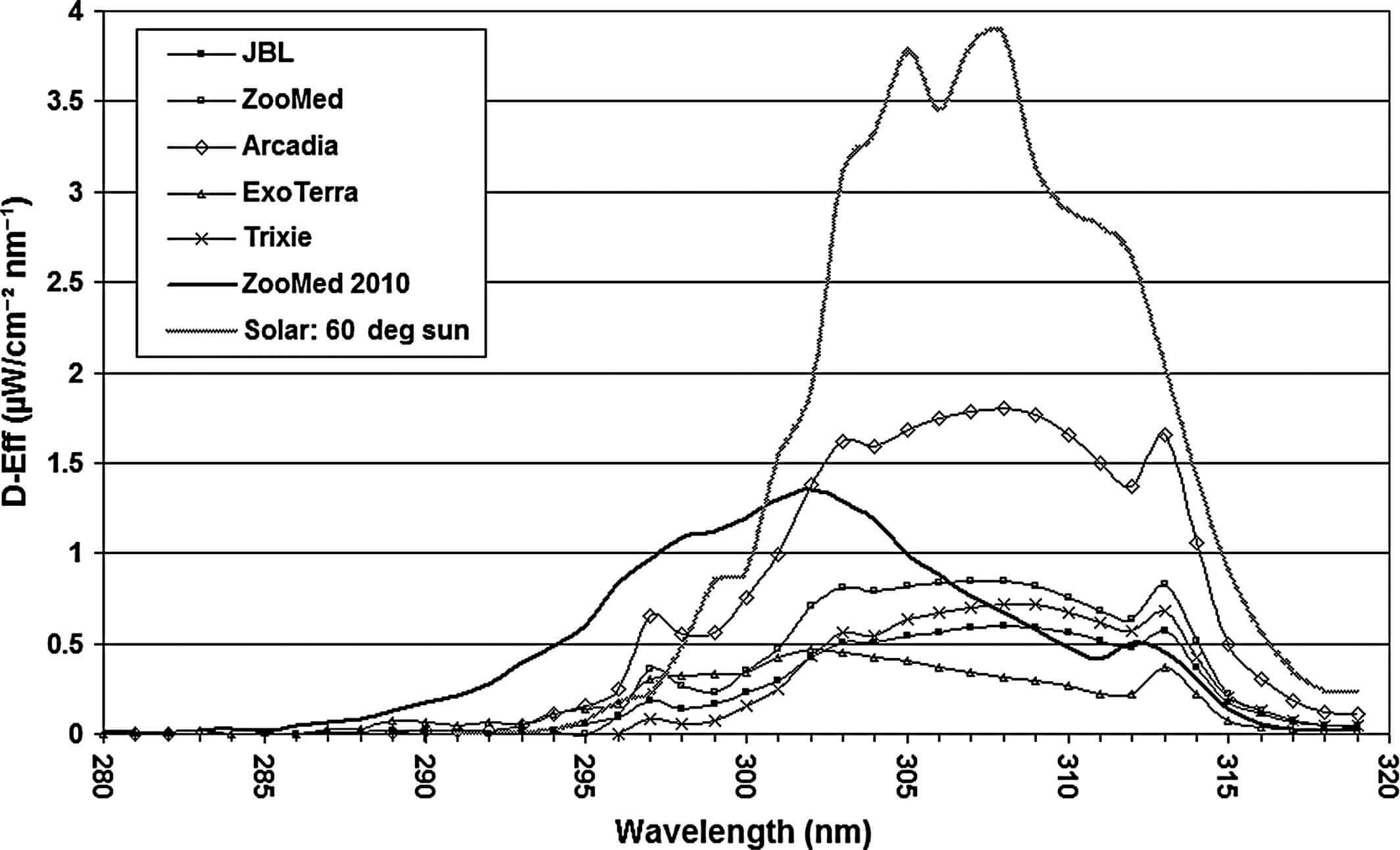
Спектры витамин D-синтезирующего излучения (D-Eff) шести компактных УФ-ламп и эталонный солнечный спектр при высоте солнца 60° (в середине утра и в середине дня летом) в Северном Квинсленде, Австралия (Bernhard et al., 1997). Расчет основан на Bouillon et al. (2006)
3.3 Концентрация в плазме крови
Ни у одного из животных в контрольной группе не было уровня 25-(ОН)D3 выше предела обнаружения, тогда как в группе Arcadia все животные имели уровни выше предела обнаружения 4нмоль/л (таблица 3). Эта группа также имела более высокую среднюю концентрацию 25-(OH)D3 в плазме, чем четыре других экспериментальных группы, из которых все были близки к пределу обнаружения. У живущих в природе бородатых агам концентрация 25-(ОН)D3 в плазме (409±56нмоль/л) в десять раз превышала концентрацию в группе Arcadia (40,00±12,85нмоль/л). Концентрация 1,25-(OH)2D3 в плазме живущих в природе бородатых агам составила 770±458пмоль/л.
Концентрация 25-(OH)D3 в плазме, кальция (Ca), фосфора (P) и мочевой кислоты (UA) в цельной крови бородатых агам (Pogona vitticeps), облучаемых контрольной или компактной УФ-лампой в течение первых 120 дней с момента вылупления (среднее значение±стандартное отклонение)
| Исследуемый компонент | Контрольная лампа (n = 4) | Лампы UVB | |||||
|---|---|---|---|---|---|---|---|
| Trixie (n = 6) | JBL (n = 6) | Arcadia (n = 6) | ZooMed (n = 4) | ExoTerra (n = 6) | p‐value | ||
| 25(OH)D3 (nmol/L) | 2.00 ± 0.00a | 4.33 ± 5.72a | 4.50 ± 3.89a | 41.00 ± 12.85b | 5.50 ± 2.65a | 10.33 ± 17.64a | .004 |
| Ca (mmol/L) | 2.20 ± 0.28a | 3.45 ± 0.37a ,b | 3.36 ± 0.41a ,b | 4.73 ± 2.69b | 3.30 ± 0.22a ,b | 3.20 ± 0.21a ,b | .027 |
| P (mmol/L) | 4.05 ± 0.22b | 3.36 ± 0.72a ,b | 2.96 ± 0.60 a ,b | 2.57 ± 0.61a | 3.57 ± 0.72a ,b | 3.00 ± 0.34a ,b | .029 |
| UA (μmol/L) | 228 ± 97a ,b | 336 ± 73a ,b | 307 ± 69a ,b | 517 ± 158b | 226 ± 98a ,b | 236 ± 98a | .012 |
a,bРазличные надстрочные буквы в строках указывают на существенные различия между группами.
Концентрации кальция в группе Arcadia были выше по сравнению с контрольной (р = 0,011), тогда как уровни Р в плазме в группе Arcadia были ниже (р = 0,026), чем в контрольной группе, в других группах значения были промежуточными. Концентрации мочевой кислоты в группе Arcadia были выше, чем в группе ExoTerra (р = 0,025).
4. Обсуждение
В этом испытании только одна лампа (лампа Arcadia) подняла уровень 25-(OH)D3 в сыворотке бородатых агам по сравнению с контролем. Эта лампа использовалась в течение двух часов в день и имела максимальную интенсивность излучения 57мкВт/см2 и UVI 1,9 (на расстоянии 25см). Эта общая доза (облучение×время) привела только к уровню 25-(ОН)D3 в сыворотке 41,00 ± 12,85нмоль/л, что составляет менее четверти концентрации, установленной ранее для растущих бородатых агам, облучаемых аналогичным образом компактной лампой ZooMed 2010 (178,4 ± 9,0нмоль/л; Oonincx et al., 2010), и только одну десятую от установленной у диких бородатых агам в их естественной среде обитания (409 ± 56нмоль/л). Максимальное излучение, доступное для агам от других ламп в испытании, было ниже (диапазон 10–27 мкВт/см2 и UVI 0,5–0,9 на расстоянии 25см), что привело к низким концентрациям 25-(OH)D3 в сыворотке. Для каждой марки во время эксперимента использовалась только одна лампа. Следовательно, это не позволяет оценить эффективность брендов, а только эффективность фактически используемой лампы и спектра света, излучаемого этой лампой.
Высокие концентрации 25-(OH)D3, обнаруженные у живущих в природе бородатых агам, вероятно, объясняются тем, что они получают гораздо большую дозу UVB от естественного солнечного света. Максимальное значение солнечного UVB-излучения, зарегистрированного в месте нахождения одной из агам, составило 506мкВт/см2 в 14:00 (рис. 4), что на порядок выше, чем у бородатых агам на самом высоком уровне (Arcadia) в испытании. Каждое зафиксированное значение между 10:00 и 15:30 превышало 100мкВт/см2. Это указывает на то, что есть сильное воздействие UVB-излучения, но мы не контролировали время воздействия, поэтому их суточная доза была неизвестна. Баскингующие виды, как правило, не остаются на долгое время полностью под воздействием облучения (Ferguson, Gehrmann, Brinker, & Kroh, 2014 , 2015 ; Ferguson et al.,2010 ). Тем не менее, UVB был доступен для живущих в природе бородатых агам в течение всего дня.
Спектральное распределение мощности источника UVB – тоже очень важный фактор, который влияет на синтез витамина D3 в коже и в итоге на концентрацию 25-(OH)D3 (MacLaughlin et al., 1982). У коротковолнового UVB-излучения более высокий потенциал синтеза витамина D3 на микроватт по сравнению с более длинными волнами, при этом в естественных условиях оптимальное значение составляет 298 нм. Более длинные волны до 320нм также запускают переход из 7-DHC в пре-витамин D3, хотя чем длиннее волна, тем выше должна быть интенсивность излучения для получения того же результата (Bernhard et al., 1997).
Спектральное распределение мощности (SPD) естественного солнечного света с началом от 290 нм до 295 нм (рис. 5) таково, что, хотя длина волны 298 нм является наиболее эффективной для синтеза витамина D3, на более длинных волнах создается гораздо более сильное излучение. В результате, наиболее продуктивная длина волны солнечного света для формирования пре-витамина D3 при высоте солнца 60° составляет 308 нм (рис. 6).
Относительное SPD всех ламп в текущем исследовании аналогично естественному солнечному свету, поэтому доза UVB-излучения, по-видимому, является основным определяющим фактором для сравнительных различий в уровне витамина D между группами в этом испытании и солнечным светом. Однако SPD компактной лампы ZooMed 2010 отличается, что приводит к другому распределению D-Eff (Рисунок 6). Эта лампа имеет большую долю коротковолнового UVB по сравнению с другими тестируемыми лампами и солнечным спектром и включает в себя высокоэнергетические длины волн <290нм, которых нет в естественном солнечном свете. Такое SPD дает наиболее эффективную длину волны для синтеза пре-витамина D3 при 302 нм. В исследовании 2010 года бородатые агамы подвергались умеренному воздействию UVB-излучения этой компактной лампы (34 мкВт/см2 UVB на расстоянии 19см, 20,7 мкВт/см2 D-Eff на расстоянии 10см), но это привело к гораздо более высоким уровням сывороточного 25-(OH)D3, чем при использовании лампы Arcadia, несмотря на более высокое общее излучение лампы Arcadia (57 мкВт/см2 на расстоянии 25см) и уровень D-Eff (28,3 мкВт/см2 D-Eff на расстоянии 10см). К сожалению, благодаря исследованиям стало понятно, что качество ламп ZooMed значительно снизилось. Почему? Это вопросы к производителю. Видимо глобализация делает свое коварное дело.
MacLaughlin et al. (1982) продемонстрировали, что характер спектра УФ-излучения определяет процент превращения 7‐DHC в различные фотопродукты – пре-витамин D3, люмистерол-3 и тахистерол-3, что дает возможное объяснение этой аномалии, требующей дальнейших исследований. Именно квазиравновесие между этими фотопродуктами и 7-DHC под воздействием УФ-излучения отвечает за саморегуляцию синтеза витамина D3, предотвращающую перепроизводство. Вполне возможно, что использование ламп со спектрами, которые сильно отличаются от солнечного света и, в частности, тех, которые излучают не характерный для Земли коротковолновый UVB (<290 нм), может каким-то образом повлиять на саморегуляцию. Очень сильно повышенные концентрации сывороточного 25-(OH)D3 были зарегистрированы у зеленых игуан, получавших только низкоуровневое облучение от тестируемой лампы этого типа (Bernard, 1995). Необходимы дополнительные исследования, чтобы определить, могут ли определенные спектры привести к перепроизводству витамина D3. Тем не менее, лампы, выдающие большую часть своего UVB-излучения на этих коротких длинах волн, оказались вредными для рептилий, вызывая повреждение глаз и кожи и смерть, и должны рассматриваться как опасные (Hibma, 2004.; Gardiner, Baines & Pandher, 2009; Бейнс, 2010).
Различные методы анализа на содержание 25-(OH)D3 были использованы для образцов крови агам в исследовании 2010 года, живущих в природе агам и агам в исследовании ламп. Было отмечено, что коммерчески доступные тесты для 25-(OH)D3 давали разные результаты не только в рамках различных методов анализа, но также между различными лабораториями, использующими одни и те же тесты (Binkley et al., 2004; Roth, Schmidt-Gayk, Weber & Niederau, 2008). Мы провели небольшое исследование (n = 20), чтобы сравнить наши два метода анализа на содержание 25-(OH)D3, используя плазму бородатых агам согласно Oonincx et al. (2013). Мы нашли значительную корреляцию между двумя методами (р = 0,006; Pearson R = 0,60), это свидетельствует о том, что результаты обоих методов можно качественно сравнивать. Однако требуется больше работы для количественной оценки потенциальных различий между образцами, анализируемыми с помощью анализа связывания и метода ID-XLC-MS/MS.
В группе Arcadia уровни кальция были выше, а уровни фосфора ниже по сравнению с контрольной группой. Кроме того, при исследовании Arcadia соотношение Ca:P, как правило, было выше по сравнению с контролем (р = 0,067). Фактически, в контрольной группе концентрация Ca была даже ниже, чем концентрация P. В этой группе у трех животных были клинические симптомы дефицита витамина D, тогда как в других группах этого не наблюдалось. Это указывает на то, что даже минимального уровня UVB-излучения достаточно для предотвращения клинических симптомов дефицита витамина D, несмотря на низкий уровень 25-(ОН)D3. У двух животных, выведенных из контрольной группы из-за клинических симптомов дефицита витамина D (тетания), образцы крови были взяты на 91-й день. Затем этих животных поместили вместе и облучали под УФ-лампой Zoomed Reptisun Desert 10,0 26 Вт, которая использовалась в одной из групп, по два часа в день в течение 3 недель. В течение первых 4 дней эти две бородатые агамы активно загорали под УФ-лампой; они вытягивались и располагались перпендикулярно лампе, таким образом максимально увеличивая облучаемую поверхность тела. После 3 недель облучения клинические симптомы исчезли, и был получен второй образец крови. Уровень 25-(OH)D3 в плазме увеличился у одной из особей (с <4нмоль/л до 26нмоль/л), тогда как у другой особи концентрация 25-(OH)D3 в плазме оставалась ниже предела обнаружения. Бородатые агамы, которые остались в контрольной группе, показали более медленный рост по сравнению с группами, которые получали облучение ультрафиолетом B. Это ожидаемо, поскольку витамин D3 и его метаболиты способствуют росту костей (van Leeuwen, van den Bemd, van Driel, Buurman, Pols, 2001). Точно так же Oonincx et al. (2010) описали задержку роста у бородатых агам, не получавших облучение, по сравнению с облучаемыми животными. Однако в этом исследовании это было очевидно только у самок бородатых агам.
Концентрации мочевой кислоты у бородатых агам в группе Arcadia были выше, чем в группе с ультрафиолетовыми лампами ExoTerra (p = .012), и, как правило, были выше, чем в контрольной группе и группе с ультрафиолетовыми лампами ZooMed (p = .075 для обеих). Хотя высокие концентрации мочевой кислоты были связаны с почечной недостаточностью (Divers & Mader, 2005), уровни, установленные в этом исследовании, находятся в пределах контрольных значений (119–595 мкмоль/л (Diethelm, Stein, & Mader, 2006; Tamukai, Takami, Akabane, Kanazawa & Une, 2011)). Будущие исследования, посвященные влиянию UVB-излучения на уровень метаболитов витамина D, должны не только определять спектральный выход ультрафиолетовых ламп, но и определять фактическое воздействие на отдельных животных. Индивидуальное размещение в сочетании с поведенческим анализом будет способствовать этому, в то же время предупреждая взаимовлияние собранных данных. Кроме того, для стандартизации UVB-излучения предпочтительнее использовать лампы с более широким пространственным диапазоном по сравнению с компактными лампами. В исследованиях, направленных на изучение метаболизма кальция, нужно использовать значения гормона околощитовидной железы, если будут разработаны анализы сыворотки рептилий.
В заключение, ни одна из ламп, использованных в этом исследовании, не подняла концентрацию 25-(OH)D3 до уровней, обнаруженных у агам, живущих в природе. Хотя, по-видимому, для предотвращения клинических симптомов было достаточно низкой дозы UVB-излучения всех ламп, субклинический дефицит витамина D может оказывать иное влияние на здоровье и благополучие бородатых агам. В этом исследовании использовались лампы с очень небольшим UVB-излучением при 295 нм. При использовании ламп с таким спектральным распределением мощности UVB-излучение, по-видимому, определяет концентрацию 25-(ОН)D3 у растущих бородатых агам. Тем не менее, значительное увеличение было видно только под лампой Arcadia с самой высокой мощностью. Следовательно, если использовать лампы с таким спектральным распределением мощности для увеличения уровня витамина D, на уровне животного может потребоваться UVB-излучение как минимум такой мощности. Компактная лампа ZooMed, использованная в предыдущем исследовании за 2010 год, с низкой интенсивностью излучения, но выдающая большой процент UVB с длиной волны ниже 295 нм, привела к значительному повышению уровня 25-(OH)D3 в плазме. Это указывает на то, что спектральное распределение мощности играет ключевую роль в синтезе витамина D, и это требует дальнейшего изучения. К сожалению, ультрафиолетовые лампы ZooMed, JBL, ExoTerra, Trixie показывают низкую эффективность работы UVB, по сравнению с лампами Arcadia. Видимо все-таки не зря, лампы Arcadia были и остаются одними из самых лучшими.
Благодарность
Авторы благодарят сотрудников исследовательского центра животных «Carus» при Вагенингенском университете за помощь в уходе за животными. А также выражают особую признательность доктору Эверт-Ян Баккеру (Dr Evert‐Jan Bakker) за советы относительно статистического анализа.
Acierno, M. J., Mitchell, M. A., Roundtree, M. K., & Zachariah, T. T.(2006). Effects of ultraviolet radiation on 25‐hydroxyvitamin D3 synthesis in red‐eared slider turtles (Trachemys scripta elegans). American Journal of Veterinary Research, 67, 2046 – 2049 .
Acierno, M. J., Mitchell, M. A., Zachariah, T. T., Roundtree, M. K., Kirchgessner, M. S., & Sanchez‐Migallon Guzman, D.( 2008). Effects of ultraviolet radiation on plasma 25‐hydroxyvitamin D3 concentrations in corn snakes (Elaphe guttata). American Journal of Veterinary Research, 69, 294-297 .
Ajibade, D., Benn, B. S., & Christakos, S.(2010). Mechanism of action of 1, 25‐dihydroxyvitamin D3 on intestinal calcium absorption and renal calcium transport. In M. F. Holick (Ed.), Vitamin D (pp. 175– 187). New York: Humana Press.
Allen, M. E., Oftedal, O. T., & Horst, R. L.(1995). Remarkable differences in the response to dietary vitamin D among species of reptiles and primates: Is ultraviolet B light essential? In M. F. Holick, & E. G. Jung (Eds.), Biological effects of light 1995 (pp. 13– 18). USA: Walter de Gruyter, Atlanta.
Aucone, B. M., Gehrmann, W. H., Ferguson, G. W., Chen, T., & Holick, M. F.(2003). Comparison of two artificial ultraviolet light sources used for Chuckwalla, Sauromalus obesus, husbandry. Journal of Herpetological Medicine and Surgery, 13, 14-17 .
Baines, F. M.(2010). Photo‐kerato‐conjunctivitis in reptiles. In S. Öfner & F. Weinzierl (Eds.), Proc. ARAV 1st Int Conference on Reptile and Amphibian Medicine, Munich, Mar 4–7 March 2010. (pp. 141– 145). München: Verlag Dr. Hut.
Bernard, J. B.(1995). Spectral irradiance of fluorescent lamps and their efficacy for promoting vitamin D synthesis in herbivorous reptiles. Comparative Nutrition Group, Dept. of Animal Science, Michigan State University.
Bernard, J. B., Allen, M. E., & Ullrey, D. E.(1997). Feeding captive insectivorous animals: Nutritional aspects of insects as food. Nutrition Advisory Group Handbook, Fact Sheet, 3, 1-7 .
Bernhard, G., Mayer, B., Seckmeyer, G., & Moise, A.(1997). Measurements of spectral solar UV irradiance in tropical‐Australia. Journal of Geophysical Research: Atmospheres (1984–2012), 102 , 8719 – 8730 .
Binkley, N., Krueger, D., Cowgill, C. S., Plum, L., Lake, E., Hansen, K. E., & Drezner, M. K.( 2004 ). Assay variation confounds the diagnosis of hypovitaminosis D: A call for standardization. Journal of Clinical Endocrinology & Metabolism, 89 , 3152-3157 .
Björn, L. O.(2008 ). Vitamin D: Photobiological and ecological aspects. New York: Springer.
Bouillon, R., Eisman, J., Garabedian, M., Holick, M., Kleinschmidt, J., Suda, T., … Webb, A.( 2006 ). Action spectrum for the production of previtamin D3 in human skin. Commission Internationale de L’Eclairage, 174, 2006 .
Carman, E. N., Ferguson, G. W., Gehrmann, W. H., Chen, T. C., & Holick, M. F.(2000). Photobiosynthetic opportunity and ability for UV‐B generated vitamin D synthesis in free‐living house geckos (Hemidactylus turcicus) and Texas spiny lizards (Sceloporus olivaceous). Copeia, 2000, 245-250 .
Diethelm, G., Stein, G., & Mader, D. (2006). Hematologic and blood chemistry values in reptiles. D. R. Mader (Ed.), Reptile medicine and surgery (pp. 1103– 1118). Philadelphia, PA: Saunders.
Divers, S.J., & Mader, D.R. (2005). Reptile medicine and surgery. Philadelphia, PA: Elsevier Health Sciences.
Ferguson, G. W., Brinker, A. M., Gehrmann, W. H., Bucklin, S. E., Baines, F. M., & Mackin, S. J.( 2010). Voluntary exposure of some western‐hemisphere snake and lizard species to ultraviolet‐B radiation in the field: How much ultraviolet‐B should a lizard or snake receive in captivity? Zoo Biology, 29, 317-334 .
Ferguson, G. W., Gehrmann, W. H., Brinker, A. M., & Kroh, G. C.(2014). Daily and seasonal patterns of natural ultraviolet light exposure of the western sagebrush lizard (Sceloporus graciosus gracilis) and the dunes sagebrush lizard (Sceloporus arenicolus). Herpetology, 70, 56-68 .
Ferguson, G. W., Gehrmann, W. H., Brinker, A. M., & Kroh, G. C.(2015). Natural ultraviolet‐b exposure of the Texas Horned Lizard (Phrynosoma cornutum) at a North Texas Wildlife Refuge. The Southwestern Naturalist, 60, 231 – 239 .
Ferguson, G. W., Gehrmann, W. H., Karsten, K. B., Hammack, S. H., McRae, M., Chen, T. C., … Holick, M. F.(2003). Do panther chameleons bask to regulate endogenous vitamin D3? Physiological and Biochemical Zoology, 76, 52-59 .
Ferguson, G. W., Gehrmann, W. H., Karsten, K. B., Landwer, A. J., Carman, E. N., Chen, T. C., & Holick, M. F.( 2005). Ultraviolet exposure and vitamin D synthesis in a sun‐dwelling and a shade‐dwelling species of Anolis: Are there adaptations for lower ultraviolet B and dietary vitamin D3 availability in the shade? Physiological and Biochemical Zoology, 78, 193-200 .
Ferguson, G., Gehrmann, W., Peavy, B., Painter, C., Hartdegen, R., Chen, T., … Pinder, J. III( 2009 ). Restoring vitamin D in monitor lizards: Exploring the efficacy of dietary and UVB sources. Journal of Herpetological Medicine and Surgery, 19, 81-88 .
Fraser, D. R.(1995). Vitamin D. Lancet, 345, 104-107 .
Gardiner, D. W., Baines, F. M., & Pandher, K.(2009 ). Photodermatitis and photokeratoconjunctivitis in a ball python (Python regius) and a blue‐tongue skink (Tiliqua spp.). Journal of Zoo and Wildlife Medicine, 40, 757-766 .
Gillespie, D., Frye, F. L., Stockham, S. L., & Fredeking, T.(2000). Blood values in wild and captive Komodo dragons (Varanus komodoensis). Zoo Biology, 19, 495-509 .
Haxhiu, D., Hoby, S., Wenker, C., Boos, A., Kowalewski, M., Lewis, F., & Liesegang, A.( 2014). Influence of feeding and UVB exposition on the absorption mechanisms of calcium in the gastrointestinal tract of veiled chameleons (Chamaeleo calyptratus) . Journal of Animal Physiology and Animal Nutrition, 98, 1021-1030 .
Heijboer, A. C., Blankenstein, M. A., Kema, I. P., & Buijs, M. M.( 2012). Accuracy of 6 routine 25‐hydroxyvitamin D assays: Influence of vitamin D binding protein concentration Clinical Chemistry, 58, 543-548 .
Hibma, J. C.(2004). Dietary vitamin D3 and UV‐B exposure effects on green iguana growth rate: Is full‐spectrum lighting necessary? Bulletin of the Chicago Herpetological Society, 39 , 145-150
Hoby, S., Wenker, C., Robert, N., Jermann, T., Hartnack, S., Segner, H., … Liesegang, A.( 2010 ). Nutritional metabolic bone disease in juvenile veiled chameleons (Chamaeleo calyptratus) and its prevention. Journal of Nutrition, 140 , 1923-1931 .
Holick, M. F.(1990). The use and interpretation of assays for vitamin D and its metabolites. Journal of Nutrition, 11, 1464-1469 .
Holick, M. F., MacLaughlin, J. A., Clark, M. B., Holick, S. A., Potts, J. T., Anderson, R. R., … Elias, P.(1980). Photosynthesis of previtamin D3 in human skin and the physiologic consequences. Science, 210, 203-205 .
Holick, M. F., Tian, X. Q., & Allen, M.(1995). Evolutionary importance for the membrane enhancement of the production of vitamin D3 in the skin of poikilothermic animals. Proceedings of the National Academy of Sciences of the United States of America, 92 , 3124 – 3126 .
Hossein‐nezhad, A., & Holick, M. F.(2013 ). Vitamin D for health: A global perspective. Mayo Clinic Proceedings Elsevier, 88, 720 – 755 .
Knotek, Z., Hauptman, K., Knotkova, Z., Hajkova, P., & Tichý, F.(2002). Renal disease haemogram and plasma biochemistry in green iguana. Acta Veterinaria Brno, 71, 333 – 340 .
Kroenlein, K., Zimmerman, K., Saunders, G., & Holladay, S.(2011). Serum vitamin D levels and skeletal and general development of young bearded dragon lizards (Pogona vitticeps), under different conditions of UV‐B radiation exposure. Journal of Animal and Veterinary Advances, 10, 229-234 .
Laszlo, J.(1969). Observations on two new artificial lights for reptile displays. International Zoo Yearbook, 9, 12-13 .
van Leeuwen, J. P., van den Bemd, G. J., van Driel, M., Buurman, C. J., & Pols, H. A.( 2001). 24,25‐Dihydroxyvitamin D3 and bone metabolism. Steroids, 66, 375-380 .
MacLaughlin, J. A., Anderson, R. R., & Holick, M. F.(1982). Spectral character of sunlight modulates photosynthesis of previtamin D3 and its photoisomers in human skin. Science, 216, 1001-1003 .
Miller, H. A.(1998 ). Urinary diseases of reptiles: Pathophysiology and diagnosis. Seminars in Avian and Exotic Pet Medicine, 7, 93-103 .
Oonincx, D. G., Stevens, Y., van den Borne, J. J., van Leeuwen, J. P., & Hendriks, W. H.( 2010). Effects of vitamin D3 supplementation and UVb exposure on the growth and plasma concentration of vitamin D3 metabolites in juvenile bearded dragons (Pogona vitticeps). Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 156 , 122 – 128 .
Oonincx, D., van de Wal, M., Bosch, G., Stumpel, J., Heijboer, A., van Leeuwen, J., … Kik, M.( 2013). Blood vitamin D3 metabolite concentrations of adult female bearded dragons (Pogona vitticeps) remain stable after ceasing UVb exposure. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 165, 196-200 .
Oonincx, D., van Leeuwen, J., Hendriks, W., & van der Poel, A.(2015). The diet of free‐roaming Australian Central Bearded Dragons (Pogona vitticeps). Zoo Biology, 34, 271-277 .
Ramer, J. C., Maria, R., Reichard, T., Tolson, P. J., Chen, T. C., & Holick, M. F.(2005). Vitamin D status of wild Ricord’s iguanas (Cyclura ricordii) and captive and wild rhinoceros iguanas (Cyclura cornuta cornuta) in the Dominican Republic. Journal of Zoo and Wildlife Medicine, 36, 188-191 .
Roth, H. J., Schmidt‐Gayk, H., Weber, H., & Niederau, C.(2008 ). Accuracy and clinical implications of seven 25‐hydroxyvitamin D methods compared with liquid chromatography–tandem mass spectrometry as a reference. Annals of Clinical Biochemistry, 45 , 153-159 .
Selleri, P., & Di Girolamo, N. (2012). Plasma 25‐hydroxyvitamin D3 concentrations in Hermann’s tortoises (Testudo hermanni) exposed to natural sunlight and two artificial ultraviolet radiation sources. American Journal of Veterinary Research, 73, 1781 – 1786 .
Tamukai, K., Takami, Y., Akabane, Y., Kanazawa, Y., & Une, Y.( 2011). Plasma biochemical reference values in clinically healthy captive bearded dragons (Pogona vitticeps) and the effects of sex and season. Veterinary Clinical Pathology, 40, 368-373 .
Wangen, K., Kirchenbaum, J., & Mitchell, M.A. (2013). Measuring 25‐hydroxy vitamin D levels in leopard geckos exposed to commercial ultraviolet B lights, Proceedings of the Association of Reptilian and Amphibian Veterinarians Conference. Indianapolis (IN), 42.
Wright, K.(2008 ). Two common disorders of captive bearded dragons (Pogona vitticeps): Nutritional secondary hyperparathyroidism and constipation. Journal of Exotic Pet Medicine, 17, 267-272 .
Number of times cited according to CrossRef: 3
Janet F. Bornman, Paul W. Barnes, T. Matthew Robson, Sharon A. Robinson, Marcel A. K. Jansen, Carlos L. Ballaré and Stephan D. Flint, Linkages between stratospheric ozone, UV radiation and climate change and their implications for terrestrial ecosystems, Photochemical & Photobiological Sciences, 10.1039/C8PP90061B, (2019).
Crossref
Jan H. Bos, Fokko C. Klip and Dennis G.A.B. Oonincx, ARTIFICIAL ULTRAVIOLET B RADIATION RAISES PLASMA 25-HYDROXYVITAMIN D 3 CONCENTRATIONS IN BURMESE PYTHONS ( PYTHON BIVITTATUS ) , Journal of Zoo and Wildlife Medicine, 10.1638/2017-0243.1, 49, 3, (810-812), (2018).
Crossref
Dennis Oonincx and Jeroen van Leeuwen, Evidence-Based Reptile Housing and Nutrition, Veterinary Clinics of North America: Exotic Animal Practice, 10.1016/j.cvex.2017.04.004, 20, 3, (885-898), (2017).
Crossref